Мембраны биологические (лат. membrana кожица, оболочка) — функционально активные поверхностные структуры клеток толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство внутриклеточных структур, а также образующие единую внутриклеточную систему канальцев, складок и замкнутых полостей.
Мембрану, ограничивающую цитоплазму клетки снаружи, называют плазматической или цитоплазматической мембраной, оболочкой клетки или плазмолеммой. Название внутриклеточных (субклеточных) мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Напр., различают митохондриальную, ядерную и лизосомную мембраны, мембраны комплекса Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и т. д. (см. Клетка).
Толщина Мембраны биологической ок. 10 нм. однако вследствие сравнительно плотной упаковки основных молекулярных компонентов (белков и липидов) в Мембране биологической, а также большой общей площади клеточных мембран они составляют обычно более половины всей массы клетки (в пересчете на сухой вес). Функции, выполняемые Мембранами биологическими, чрезвычайно важны и разнообразны: формирование клеточных структур, поддержание внутриклеточного гомеостаза, участие в процессе возбуждения и проведения нервного импульса, фото-, механо- и хеморецепция, всасывание, секреция и газообмен, тканевое дыхание, запасание и трансформация энергии и т. д. Всем этим определяется общебиологическое значение М. б. как универсальной и доминирующей формы структурной и функциональной организации живой материи.
Мембрана: строение и функции
Огромное значение Мембран биологических определяется важностью перечисленных выше функций в процессах нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при нарушениях функций М. б. и проявляющихся на различных уровнях организации— от клетки и ее субклеточных систем до тканей, органов и организма в целом. Подавляющее число известных заболеваний человека и животных являются либо прямым следствием нарушений мембран, либо процессами и состояниями, в большей или меньшей степени сопряженными с ними.
- 1 История
- 2 Состав и структура биологических мембран
- 3 Важнейшие функции биологических мембран
- 4 Методы изучения биологических мембран
История
Термин «мембрана» для обозначения поверхностных образований клетки был введен нем. исследователями Молем (H. Mohl, 1851) при описании плазмолиза клеток растений и Негели (С. W. Nageli, 1855) при изучении механизма осмотических явлений и проникновения в клетки красителей. В 1877 г. Пфеффер (W. F. Ph.
Pfeffer) обосновал существование клеточной мембраны, продемонстрировав общность осмотических свойств клеток и осмометров, образованных искусственными полупроницаемыми мембранами. В 80-х гг. 19 в. Х. де Фрис обнаружил, что цитоплазма растительных клеток заключена между двумя мембранами — плазмолеммой и тонопластом.
Первые указания на то, что в состав мембраны клетки входят липиды, получил Овертон (Е. Overton, 1895—1902), к-рый обнаружил прямую связь между растворимостью многих веществ в липидах и их скоростью проникновения в клетку. В 1925 г. Гортер (E. Gorter) и Гренделл (F. Grendell) экспериментально показали, что в мембране эритроцитов количество липидов достаточно для построения двойного непрерывного слоя.
Жидкостно-мозаичная модель структуры клеточной мембраны (видео 1) | Мембранный транспорт | Биология
Это позволило им высказать предположение, что поверхностная мембрана эритроцита содержит бимолекулярный липидный слой. Примерно в это же время Фрик (H. Fricke) измерил электрическую емкость мембран эритроцитов и получил величину —0,81 мкф/см 2 , что соответствует слою диэлектрика (липида) толщиной 3,3 нм и толщине бимолекулярного слоя, образованного жирными к-тами с 16—17 углеродными атомами.
В 30-х гг. 20 в. Харви и Даниэлли (Е. N. Harvey, J, F, Danielli) показали, что величина поверхностного натяжения на границе жировых капель и цитоплазмы клеток ниже (~0,1 дин/см), чем для чистой границы раздела липид — вода (~10 дин/см). Это указывает на возможность спонтанного образования ассоциатов растворимых белков с ориентированными слоями липидов. Развивая эти представления, Даниэлли и Давсон (H.
Davson) в 1935 г. выдвинули первую гипотезу о строении М. б., согласно к-рой мембрана состоит из двойного липидного слоя, покрытого с двух сторон слоями глобулярных белков.
Непосредственная возможность наблюдать биол, мембраны появилась лишь в 50-х гг. 20 в. вследствие развития метода электронной микроскопии и методик приготовления ультратонких срезов. Полученные снимки мембран позволили представить М. б. как трехслойные структуры толщиной порядка 10 нм для плазматических и несколько меньшей — для субклеточных мембран. Робертсон (J.
D. Robertson) выдвинул гипотезу об однотипности строения всех биол, мембран и предложил унитарную схему строения мембраны. По Робертсону, белки М- б. могут разворачиваться на поверхности двойного липидного слоя под действием сил электростатического взаимодействия с заряженными головками молекул фосфолипидов; на наружной поверхности мембраны располагаются еще и молекулы гликопротеидов. Эта схема отражала важный принцип строения мембран — ее асимметричность.
Постепенно под влиянием новых фактов, и в первую очередь зернистости структуры мембран, к-рая просматривалась на снимках, полученных при большом увеличении, первоначальные представления о трехслойности мембран были пересмотрены. Вначале Луси (J. Lucy) высказывает предположение о мицеллярной организации липидного слоя в мембране. Шёстрандом (F.
S. Sjostrand) была выдвинута гипотеза о глобулярной организации цитоплазматической мембраны в целом. Позже Грин (D. Е. Green) предложил схему организации мембран из субъединиц и сформулировал принцип повторяющихся единиц применительно к внутренней мембране митохондрий.
Несмотря на то, что эта модель была тщательно разработана, она не давала удовлетворительного объяснения хорошо известному факту низкой проницаемости мембран для ионов. При этом тетерогенные частицы, образующие мембраны, не удалось получить при фрагментации мембран. Вызывало сомнение и то, что липидам в этой модели отводилась более чем скромная роль, а также практически полное отсутствие учета липид-белкового взаимодействия. В других моделях напр., в модели Бенсона) липид-белковому взаимодействию отводится, напротив, центральная’ роль в формировании мембран.
В дальнейшем оказалось, что отказ от представлений о существовании сплошного бимолекулярного слоя был преждевременным. По новым воззрениям белки не выстилают поверхность липидного слоя, а как бы плавают на поверхности в виде отдельных глобулярных молекул или частиц, в большей или меньшей степени погруженных в мембрану.
Эта жидкомозаичная модель, предложенная Ленардом и Сингером (J. Lenard, S. Singer), позволяет удовлетворительно объяснять целый ряд фактов, в частности зависимость многих физиологических функций мембран и активности отдельных мембранных ферментов от фазового состояния липидов в мембране и степени ее текучести (вязкости). Однако белок-белковое взаимодействие в этой модели учитывается недостаточно, и она не позволяет удовлетворительно объяснить экспериментально установленный факт сохранения структуры и основных параметров мембраны при извлечении из нее значительного количества липидов. Эти факты были в дальнейшем учтены в белковокристаллической модели, предложенной Вандеркои (G. Vanderkooi) и Грином, отличающейся фактически лишь наличием протяженных белковых структур, образующихся в результате осуществления дальнодействующих белок-белковых связей.
Наиболее популярными моделями Мембран биологических, стали жидкомозаичные. В то же время становится все более ясным, что мембраны отличаются друг от друга: они разнообразны по составу и специфичны в функциональном отношении. Тонкая организация мембран тесно взаимосвязана с их функциональным состоянием; и то, и другое характеризуется чрезвычайной чувствительностью к действию внешних факторов. При этом нек-рые мембраны проявляют отдельные черты, свойственные различным моделям, а иногда (напр., у нек-рых бактерий) мембраны представляют собой как бы набор фрагментов, соответствующих какой-то одной из разработанных моделей.
Состав и структура биологических мембран
Основные компоненты М. б., как правило, синтезируются вне мембранной системы механизмов. Их включение в мембраны еще недостаточно изучено. В наиболее простых случаях, вероятно, участвуют обменные белково-липидные комплексы (напр., фосфолипиды). Возможно и сопряжение процессов биосинтеза с быстрым включением компонентов в мембрану (напр., Na+, К+ — АТФ-азы).
По-видимому, отдельные компоненты могут встраиваться в мембраны независимо друг от друга, т. к. в мембранах, как правило, отсутствуют специфические центры роста. Вместе с тем не вызывает сомнений, что нек-рые липид-зависимые ферменты в определенных ситуациях встраиваются в мембрану одновременно с липидным окружением.
Мембранные структуры сформированы за счет сравнительно слабых сил гидрофобных и электростатических (ван-дер-ваальсовых) взаимодействий (см. Молекула, Строение вещества). Ковалентные связи в формировании мембранных структур играют второстепенную роль. В связи с этим мембраны обладают рядом особых физ.-хим. свойств.
Так, молекулярные компоненты сохраняют в мембранах довольно высокую подвижность. Различают внутримолекулярную подвижность, связанную с вращательной подвижностью вокруг одиночных связей, вращательную подвижность молекул в целом, движение молекул в плоскости мембраны (латеральная подвижность) и вертикальную подвижность молекулярных компонентов мембран (либо частичную, либо с переходом молекулы из одной половины мембраны в другую).
Обычно мембраны функционируют при температурах, когда жирнокислотные остатки фосфолипидов находятся в жидком (точнее жидкокристаллическом) состоянии (см. Жидкие кристаллы). В этом состоянии скорость диффузии фосфолипидов обеспечивает передвижение липидной молекулы за время порядка одной секунды. Переходы же липидов с одной половины бимолекулярного слоя мембраны на другую (так наз. переходы флип-флоп) совершаются сравнительно редко. Этот процесс, очевидно, должен быть заторможен в мембранах с выраженной асимметрией липидного состава, напр, в мембранах эритроцитов, в к-рых сфингомиелин и фосфатидилхолин находятся главным образом в наружной половине мембраны, а фосфатидилэтаноламин и фосфатидилсерин — на ее внутренней стороне.
Белковые молекулы также проявляют довольно большую свободу движения в мембране. Так, скорость вращательного движения и латеральной подвижности нек-рых гликопротеидов (антигенных и рецепторных белков) соответствует скорости их свободной диффузии в среде соответствующей вязкости.
Обнаружена также вертикальная подвижность мембранных белков, обычно сопряженная с их функциональным состоянием. Напр., глубина погружения родопсина в мембрану меняется в зависимости от функционального состояния белка. Свободное движение, однако, присуще не всем мембранным белкам. Часто они образуют устойчивые плотноупакованные образования (область межклеточных контактов, бляшки пурпурных мембран Halobacterium и др.) или строго ориентированные системы (компоненты электрон-транспортных цепей митохондрий, хлоропластов, эндоплазматического ретикулума и др.). Очевидно, подвижность белков ограничена в тех случаях, когда они находятся в липидном микроокружении, отличном по составу от основной массы липидов мембран.
В соответствии с различными типами подвижности мембранных компонентов наблюдается значительная гетерогенность в вязкости различных участков М. б. Низкие значения вязкости наблюдаются в углеводородном слое липидов, причем вязкость убывает к середине слоя в соответствии с градиентом вращательной подвижности углеводородных цепей липидов. Минимальные значения вязкости в середине липидного би-слоя, измеряемой при температурах выше точки фазового перехода, составляют сотые доли пуаз. Вязкость полярного слоя мембраны, измеренная по вращательной и латеральной подвижности молекул, соответствует единицам пуаз. В то же время общая вязкость мембран как структурного образования клетки, измеренная по величине механической деформации мембраны в целом, достигает 10 7 — 10 8 пуаз. Нек-рые другие физ.-хим. свойства мембран приведены в таблице.
Таблица. Сравнение некоторых физико-химических свойств биологических и искусственных фосфолипидных мембран
Биологические мембраны (t° 25°)
Искусственные фосфолипидные мембраны (t° 36°)
Источник: xn--90aw5c.xn--c1avg
Принцип работы клеточной мембраны
Клеточная мембрана – это трёхслойная липопротеиновая (жиро-белковая) оболочка, отделяющая каждую клетку от соседних клеток и окружающей среды, и осуществляющая управляемый обмен между клетками и окружающей средой.
Мембрана обеспечивает:
- Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток;
- Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала;
- Специфику межклеточных контактов.
Функции, выполняемые клеточной мембраной:
Барьерная
Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность). Вредные или неподходящие молекулы, благодаря барьерной функции клеточной мембраны, просто не могут проникнуть внутрь клетки.
Транспортная
Сквозь мембрану проходит пассивный, активный, регулируемый и избирательный обмен. Пассивный обмен подходит для жирорастворимых веществ и газов, состоящих из очень маленьких молекул. Такие вещества проникают внутрь и выходят из клетки без затрат энергии, свободно, методом диффузии. Активная транспортная функция клеточной мембраны задействуется тогда, когда в клетку или из неё нужно провести необходимые, но трудно транспортируемые вещества.
Энергетическая
Фотосинтез и клеточное дыхание были бы невозможны без участия белка, содержащегося в клеточной мембране. Именно через белковые каналы происходит важный клеточный энергообмен, в этом заключаются самые главные функции белка в клеточной мембране.
Другие функции, выполняемые клеточной мембраной:
- Матричная
- Механическая
- Защитная
- Рецепторная
- Ферментативная
- Биопотенциальная
- Маркировочная
Также через клеточную мембрану осуществляется клеточный обмен, который может проходить тремя разными реакциями:
- Фагоцитоз (встроенные в мембрану клетки-фагоциты захватывают и переваривают различные питательные вещества).
- Пиноцитоз (процесс захвата мембраной клетки, соприкасающиеся с ней молекулы жидкости. Для этого на поверхности мембраны образуются специальные усики, которые как будто окружают каплю жидкости, образуя пузырек, который впоследствии «проглатывается» мембраной).
- Экзоцитоз (представляет собой обратный процесс, когда клетка через мембрану выделяет секреторную функциональную жидкость на поверхность).
Наш адрес:
Санкт-Петербург, Каменноостровский пр. 61/2, вход в арку со стороны улицы Чапыгина
Режим работы: пн — пт: 10:00 — 18:00
Источник: mederispb.ru
1.3. Состав и строение мембран.
1.3.1. Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:

- состоят из липидов, белков и углеводов;
- являются плоскими замкнутыми структурами;
- имеют внутреннюю и внешнюю поверхности (асимметричны);
- избирательно проницаемы.
1.3.2. Схема строения биологической мембраны, представлена на рисунке 1.3. Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойством амфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду. Рисунок 1.3. Схема строения биологической мембраны (по Сингеру и Николсону). Мембранные липиды выполняют роль растворителя мембранных белков, создавая жидкую среду, в которой они могут функционировать. По степени влияния на структуру бислоя и по силе взаимодействия с ним мембранные белки делят на интегральные и периферические. Важнейшие особенности интегральных и периферических белков представлены в таблице 1. Таблица 1.1
Характеристика мембранных белков
- структурные;
- каталитические;
- рецепторные;
- транспортные.
1.4. Липидный состав мембран
1.4.1. Как уже упоминалось (1.1), компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран, представлены на рисунке 1.4. Выучите эти формулы. 
- они содержат чётное число атомов углерода (С16 – С20);
- двойная связь в ненасыщенных жирных кислотах располагается между 9 и 10 атомами углерода;
- в полиненасыщенных жирных кислотах двойные связи разделены метиленовыми группами (СН=CH-CH2-CH=CH), то есть являются несопряжёнными;
- двойные связи находятся в цис-конформации, что приводит к изгибу углеводородной цепи.
1.4.2. Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины. Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида представлена на рисунке 1.5. Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:  В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой).
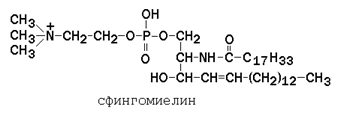
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой). 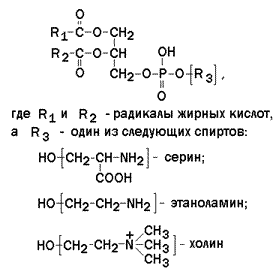 Рисунок 1.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 1.6). Соединение сфингозина и жирной кислоты получило название церамид.
Рисунок 1.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 1.6). Соединение сфингозина и жирной кислоты получило название церамид. 
 Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы: 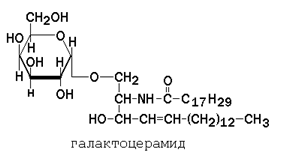 а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 1.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани.
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 1.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани. 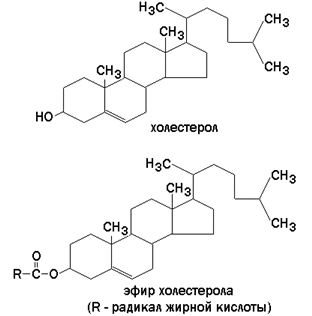 Рисунок 1.7. Структурные формулы холестерола и его эфира. 1.4.3. Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 1.4.4. Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 1.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 1.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
Рисунок 1.7. Структурные формулы холестерола и его эфира. 1.4.3. Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 1.4.4. Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 1.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 1.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран. 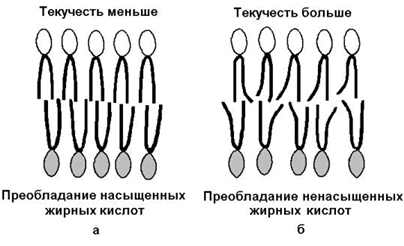 Рисунок 1.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 1.9,а).
Рисунок 1.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 1.9,а).  Рисунок 1.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 1.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул.
Рисунок 1.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 1.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул.
Источник: studfile.net
Что входит в состав мембраны
Общее строение клетки: ядро, цитоплазматическая мембрана, цитоплазма
Клетка — основная функциональная единица организма. Ядро клетки служит хранилищем огромного объёма генетической информации и одновременно центром её активной экспрессии. Существует большое количество различных типов клеток (клетки эпителия, печени, нервных волокон и др.), особенности метаболизма которых обусловлены находящимися в их цитоплазме органеллами, а также множеством растворимых ферментов, характерных для каждого вида клеток.
Цитоплазматическая мембрана, или плазмолемма, — барьер для растворимых в воде молекул, который отделяет внутреннее содержимое клетки от внешней среды. Она состоит из двух параллельных рядов фосфолипидов, которые образуют гидрофобную липидную прослойку между двумя гидрофильными слоями из фосфатных групп.
Плазмолемма пронизана различными белками, гидрофобные части которых находятся внутри билипидного слоя, а гидрофильные — на внешней и внутренней поверхности мембра ны. Микроворсинки — удлинения на верхней (апикальной) части плазмолеммы, которые увеличивают поверхность мембраны и облегчают обмен молекулами.
Ядро клетки. Генетическая информация заключена в хромосомах, которые находятся в ядерном матриксе. Матрикс — сетчатый внутриядерный каркас, состоящий из белкового материала и тесно примыкающий к ядерной оболочке.
Ядрышком называют морфологически выраженную структуру внутри ядра, в которой происходит синтез рибосомальной РНК (рРНК). В ядре клеток человека обычно присутствует одно ядрышко, в котором во время интерфазы возникают ядрышковые организаторы акроцентрических хромосом.
Ядро окружено двойной мембраной, называемой ядерной оболочкой, которая пронизана ядерными порами.
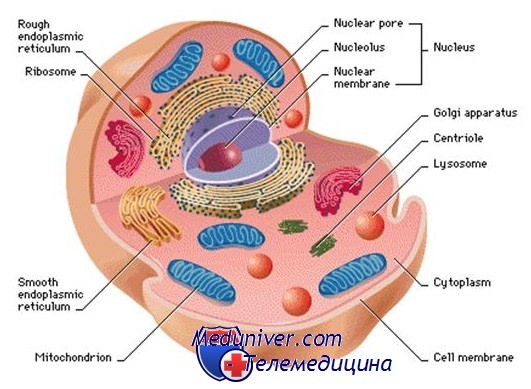
Цитоплазма клетки. Цитоплазма состоит из гелеобразного цитозоля, содержащего запасы гликогена, липидные вкрапления и свободные рибосомы, который пронизан рядами взаимосоединённых волокон и трубочек, образующих цитоскелет. Основные структурные компоненты цитоскелета — микротрубочки, микрофиламенты и промежуточные филаменты.
Микротрубочки — прямые полые цилиндры, стенки которых состоят из чередующихся молекул а- и b-тубулина. Они исходят из клеточного центра (центросомы), который имеет пару центриолей— цилиндрических структур, образованных девятью триплетами микротрубочек. Подобное строение свойственно также базальным тельцам реснитчатого эпителия.
Сеть микротрубочек играет важную роль в поддержании структуры и размера клетки, а также при расхождении хромосом во время деления и движения ресничек и сперматозоидов.
Микрофиламенты представляют собой двуспиральные полимеры белка актина и находятся в основном по периметру клетки. Они участвуют в движении клетки и изменении её формы.
Промежуточные филаменты имеют трубчатую структуру и соединяют десмосомы. В зависимости от вида клетки в их состав входит один или несколько из пяти определённых белков.
Митохондрии — самые крупные и наиболее распространённые в цитоплазме органеллы, основной функцией которых служит обеспечение организма энергией посредством синтеза АТФ. Митохондрии — самовоспроизводящиеся полуавтономные органеллы, содержащие рибосомы и до десяти и более копий кольцевых нитей митохондриальной ДНК.
Данная ДНК кодирует митохондриальные гены. В митохондриях присутствуют ферменты, необходимые для функционирования цикла трикарбоновых кислот, а также большое количество ферментов, участвующих в окислении жирных кислот.
Пероксисомы частично отвечают за детоксикацию различных веществ (в том числе этанола), однако их основная задача — окисление жирных кислот.
Эндоплазматическая сеть (ЭПС) — основной центр синтеза белков и липидов, который также служит начальным этапом секреторного пути белков. ЭПС представляет собой обширный лабиринт из связанных с мембраной каналов, который соединяется непосредственно с ядерной оболочкой.
Вблизи ядра на поверхности ЭПС есть рибосомы (гранулярная ЭПС), в то время как на участках, расположенных дальше, рибосомы отсутствуют (агранулярная или гладкая ЭПС). ЭПС играет важную роль в нейтрализации токсинов. Белки, синтезируемые в ней, затем попадают в комплекс Гольджи — ряд расположенных друг над другом сплюснутых везикул. После этого белки депонируются или попадают в секреторные везикулы для осуществления экзоцитоза, т.е. выведения из клетки в ответ на внешнее воздействие.
Эндоцитоз. Эндоцитозом называют процесс поглощения и переработки клеткой компонентов окружающей среды. При опосредованном рецепторами пиноцитозе происходит захват мелких частиц путём образования везикулы с жидкостью на поверхности цитоплазматической мембраны и её последующего поглощения клеткой. При этом образуются окаймлённые впячивания. Более крупные частицы связываются с мембраной и поглощаются в составе фагоцитарных вакуолей (фаголизосом); растворы поглощаются при помощи жидкостного пиноцитоза.
Содержимое пиноцитарных и фагоцитарных везикул, которые часто называют эндосомами, обычно обрабатывают лизосомы, содержащие разрушающие ферменты — лизоцимы.
Межклеточные соединения. В случае плотного соединения образуется непроницаемая перемычка между внешней (апикальной) и базолатеральной поверхностями эпителиальных клеток. При липких соединениях клетки связаны с помощью опоясывающих (длинные волокна) и точечных (расположены непосредственно в месте скрепления) десмосом. Гемидесмосомы (полудесмо-сомы) соединяют эпителиальные клетки через базальные мембраны (производные экстрацеллюлярного матрикса).
Щелевые соединения (нексусы) возникают в комплексах соединённых клеток. При этом через поры (щели) возможно сообщение между соседними клетками.
Недостаточность функций лизоцимов — причина некоторых наследственных заболеваний, таких, как, например, болезни Тея—Сакса (ранняя детская амавротическая идиотия), Фабри (наследственный дистопический липоидоз) и Гоше (наследственный глюкоцереброзидоз). В результате нарушения процесса поглощения клеткой липопротеинов возникает наследственная гиперхолестеринемия. При синдроме Цельвегера, для которого характерны деформации лица, снижение мышечного тонуса, увеличение печени и кисты почек, отсутствуют пероксисомы.
Причина болезни Шарко—Мари—Тута, сцепленной с Х-хромосомой, — дефект белка, участвующего в щелевом соединении клеток.
Большинство лекарственных препаратов вступают во взаимодействие с рецепторами цитоплазматической мембраны. Различные противоопухолевые препараты, такие, как винкристин или винбластин, повреждают систему микротрубочек, в то время как колхицин, применяемый для исследования хромосом, угнетает клетки во время метафа-зы митоза. Клофибрат снижает продукцию дополнительных пероксисом, его используют для снижения уровня липопротеинов в сыворотке крови.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Значение современной генетики в медицине. Задачи
- Общее строение клетки: ядро, цитоплазматическая мембрана, цитоплазма
- Строение хромосом: хроматин, центромера, теломера
- Структура ДНК: центромеры, теломеры
- Репликация ДНК. Механизмы
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Источник: meduniver.com
Химический состав мембран
Еще в 30-х годах ХХ века наличие дифференцированной мембранной структуры на поверхности клеток представлялось далеко не бесспорным. Прямые морфологические доказательства наличия мембраны были немногочисленны или отсутствовали. Первые указания на свойства клеточной поверхности, лимитирующей диффузию красителя внутрь клетки, сделал в середине XIX века К.В.Нагели. Он также показал, что клетки проявляют осмотические свойства: набухают в разбавленных растворах и сжимаются в концентрированных. Морфологические данные о существовании клеточной мембраны были получены только после разработки методов приготовления ультратонких срезов тканей.
В настоящее время не вызывает сомнений, что биологические мембраны образуют наружную оболочку всех живых клеток, а также участвуют в формировании многочисленных внутриклеточных органелл.
Мембраны выполняют многочисленные функции, обеспечивая целостность клеток и тканей и их активность.
Основные функции (выполняются всеми без исключения мембранами):
1. Компартментализация – участие мембран в образовании изолированных отсеков (компартментов). Мембраны всегда образуют замкнутые структуры, самая крупная из них – цитоплазматическая мембрана.
3. Рецепторная (сигнальная) функция – связывание мембранными рецепторами внеклеточных эффекторов – биологически (физиологически) активных веществ, что приводит к изменению активности регуляторных белков клетки.
4. Ферментативная функция обеспечивается мембранно-связанными белками – ферментами, которые образуют своеобразные конвейеры.
5. Факультативные функции, присущие не всем мембранам:
· Транспорт электронов и фосфорилирование в дыхательной цепи (способны выполнять мембраны митохондрий, хлоропластов).
· Преобразование внешних стимулов в электрические импульсы.
· Проведение биоэлектрических импульсов (характерны для мембран электровозбудимых клеток: нервной и мышечной ткани).
· Высвобождение синаптических нейромедиаторов и пиноцитоз.
Исследования показали, что основными химическими компонентами мембран являются липиды (30%), белки (60%) и углеводы (10%). Минорными компонентами мембран являются нуклеиновые кислоты, полиамины, неорганические ионы.
Липиды, входящие в состав мембран, принадлежат к трем классам. Это фосфолипиды, гликолипиды и стероиды.
Фосфолипидыпостроены по единому плану, их молекула имеет полярную головку и два неполярных хвоста.
В состав головки фосфолипида обязательно входят:
А) остаток спирта (реже аминокислоты);
Б) остаток фосфорной кислоты;
В) по вариантам, либо:
— многоатомный спирт сфингозин (сфингофосфолипиды).
Неполярные хвосты всех молекул фосфолипидов образуются жирными кислотами, определяющими разнообразие их представительства.
Глицерофосфолипиды мембран растительных клеток содержат в основном пальмитиновую (С16:1 (9)), олеиновую (С18:1(9)), линолевую (С18:2(9,12)) жирные кислоты. Жирные кислоты с количеством атомов углерода более 20 – 24 в растительных клетках встречаются редко.
Глицерофосфолипиды мембран животных клеток содержат пальмитиновую, олеиновую, стеариновую (С18:0) жирные кислоты, а также кислоты (арахидоновая и др.) с числом атомов углерода, превышающим 20.
Обычно из двух жирных кислот, образующих гидрофобный хвостглицерофосфолипида, одна ненасыщенная. Она образует связь со вторым атомом углерода глицерина и одна из ее двойных связей обязательно находится на уровне 9 атома углерода, считая от головки.
Остаток спирта (аминокислоты) головки глицерофосфолипида связан сложноэфирной связью с фосфорной кислотой. Глицерофосфолипид, лишенный спирта (аминокислоты), называется фосфатидной кислотой, которая является промежуточным продуктом в биосинтезе фосфолипидов и в свободном виде практически не встречается.
В зависимости от того, какой именно спирт (аминокислота) входит в состав головки молекулы, различают несколько классов глицерофосфолипидов. Самые распространенные:
· фосфатидилхолин, содержащий спирт холин;
· фосфатидилэтаноламин, содержащий спирт этаноламин;
· фосфатидилинозитол, содержащий циклический спирт инозитол;
· фосфатидилсерин, содержащий остаток гидроксиаминокислоты – серина.
Примерами сфингофосфолипидов могут служить сфингомиелины, которые в составе полярной головки содержат фосфохолин или фосфоэтаноламин. В составе сфингофосфолипидов одна жирная кислота. Роль второго неполярного хвоста выполняют углеродные структуры многоатомного спирта сфингозина.
Таким образом, фосфолипиды содержат группировки двух видов – полярные гидрофильные головки и неполярные гидрофобные хвосты. Вследствие этого фосфолипиды обладают амфипатическими свойствами. При рН = 7 остаток фосфорной кислоты в полярной головке заряжен отрицательно.
Гликолипиды – липиды, содержащие 2 остатка жирных кислот, спирт сфингозин и остатки моно- (цереброзиды) или олигосахаров (ганглиозиды).
Стероиды содержат стероидное ядро, образованное тремя гексагональными полностью насыщенными кольцами и одним циклопентановым кольцом. Стероиды представлены в основном холестерином (в животных клетках) или ситостерином и стигмастерином (в растительных клетках).
Физиологическая роль различных классов липидов
Фосфолипиды выполняют структурную функцию, образуя липидный бислой мембраны. Кроме этого, они могут выполнять и важные физиологические функции. Так, арахидоновая кислота, состоящая из цепи с 20 атомами углерода и с 4 двойными связями, входящая в состав фосфолипидов, служит предшественником таких биологически активных веществ, как простагландины.
Гликолипиды широко представлены в различных тканях, в частности, в нервной. Они локализованы преимущественно на наружной поверхности цитоплазматической мембраны, где их углеводные компоненты входят в число других углеводов клеточной поверхности. Ганглиозиды участвуют в дифференцировке нейрональной ткани.
Ганглиозиды других клеток определяют видоспецифичность и регулируют межклеточные контакты. В иммуннокомпетентных клетках они участвуют в формировании иммунной реакции. Холестерин выполняет важную роль в модификации бислоя: он, в частности, регулирует упаковку и подвижность фосфолипидов мембраны. Кроме того, холестерин служит предшественником в образовании половых гормонов, гормонов коры надпочечников, а также желчных кислот.
Липид-липидные взаимодействия. Динамика липидов в мембране
Длина растянутой углеводородной цепи жирной кислоты, состоящей из 18 атомов углерода, составляет 2 нм, на полярную головку приходится еще 0,5 – 0,7 нм, следовательно, толщина бислоя липидов должна превышать 4 нм, но в действительности толщина двойного слоя липидов оказывается равной 3,5 – 4 нм. Причиной обнаруженного феномена является рыхлое, а не упорядоченное расположения остатков жирных кислот. Раз одна из двух жирных кислот, образующих гидрофобный хвост, ненасыщенная, как правило, увеличивается вероятность образования цис-конформации.
Наличие цис-изомеров жирных кислот в составе фосфолипидов, а также высокая скорость вращения вокруг С-С связей жирно-кислотных радикалов (например, частота вращения вокруг единичной С-С связи составляет 10 10, а вокруг двойной С=С связи – 10 8 с -1) обеспечивают неупорядоченное расположение гидрофобных хвостов. Различные конфигурации молекул жирных кислот, возникающие при поворотах вокруг С-С связей, называют ротамерами или конформерами.
Процесс изменения конформации молекул за счет таких поворотов называется транс-гош-изомеризацией. Гош-конформация (“гош” – скошенный) аналогична цис-конформации. Вероятность транс-гош-перехода весьма велика и еще более возрастает при увеличении температуры. При переходе из транс- в гош-конформацию образуются складки или кинки (от англ. Kink – петля).
Такого рода изменения способны захватывать целые кластеры мембранных липидов. Считается, что именно с образованием кинков связан транспорт воды и других веществ через мембраны.
Липиды способны совершать вращательные движения вокруг оси молекулы (вращательная диффузия), перемещаться вдоль одного слоя (латеральная диффузия), а также перемещаться из одного слоя в другой (трансмембранный переход или флип-флоп). Наибольшую скорость имеют вращательная и латеральная диффузия. Например, коэффициент латеральной диффузии составляет 1,8×10 -8 см 2 с -1.
Это соответствует частоте парных перестановок соседних молекул 10 7 с -1. Наиболее медленным процессом является флип-флоп, что связано с высоким уровнем энергии, необходимой для проталкивания заряженных полярных головок через средний углеводородный слой мембраны. Поперечная диффузия молекул фосфолипидов на расстояние 5 нм занимает в 10 9 раз больше времени, чем диффузия на то же расстояние в латеральном направлении.
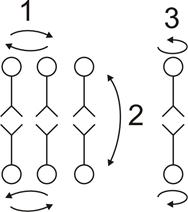 |
Рис.8. Виды диффузии липидов в мембране 1-латеральная диффузия; 2-трансмембранный переход или флип-флоп; 3-вращательная диффузия. |
Сочетание быстрой диффузии молекул липидов вдоль мембраны и медленной диффузии поперек мембраны имеет большое значение для функционирования мембран. Благодаря этому поддерживается упорядоченность в молекулярной структуре мембраны, определенная ориентация белков поперек мембраны, что имеет значение для направленного переноса веществ через мембрану, кроме того, обеспечивается асимметрия бислоя липидов. Функциональная асимметрия мембраны – один из важнейших аспектов, характеризующих липид – липидные взаимодействия. Асимметрия бислоя означает, что состав липидов каждого из слоев неодинаков.
Источник: studopedia.ru